Outline function of ANS. (Cross-reference with anatomy curriculum).
ANS as the effector component of homeostasis
Principles of neurochemical synaptic transmission
– Neurotransmitter vesicles and their role following Calcium release at the bouton
– Post-synaptic [excitatory] and pre-synaptic [inhibitory] receptors
– Role of cholinesterase, MAO and COMT in synaptic transmission
Curriculum
The ANS has sympathetic and parasympathetic (SNS and PNS) systems.
ANS preganglionic neurons originate in the CNS and synapse with non-myelinated postganglionic neurons in the peripheral ganglia. From the ganglia (postganglionic) neurons innervate the target organs.
All Preganglionic neurons(of both SNS and PNS) release acetylcholine in the synapstic cholinergic nicotinic receptors.
PNS ganglia are located close to or in the target organ (para=side by side or beside).
SNS ganglia are located in two sympathetic chains on either side of the vertebral column (paravertebral ganglia), or in diffuse prevertebral ganglia of the visceral plexuses of the abdomen and pelvis.
This is the reason why PNS postganglionics are shorter compared to SNS. An exception – SNS innervation of the adrenal gland, where preganglionic neurons directly innervate the adrenal medulla.
If an organ is innervated by both PNS and SNS, they often act antagonistically with a high degree of central coordination.
SNS – ‘flight or fight’ responses
PNS – ‘rest and digest’ responses.
Neurochemical transmission
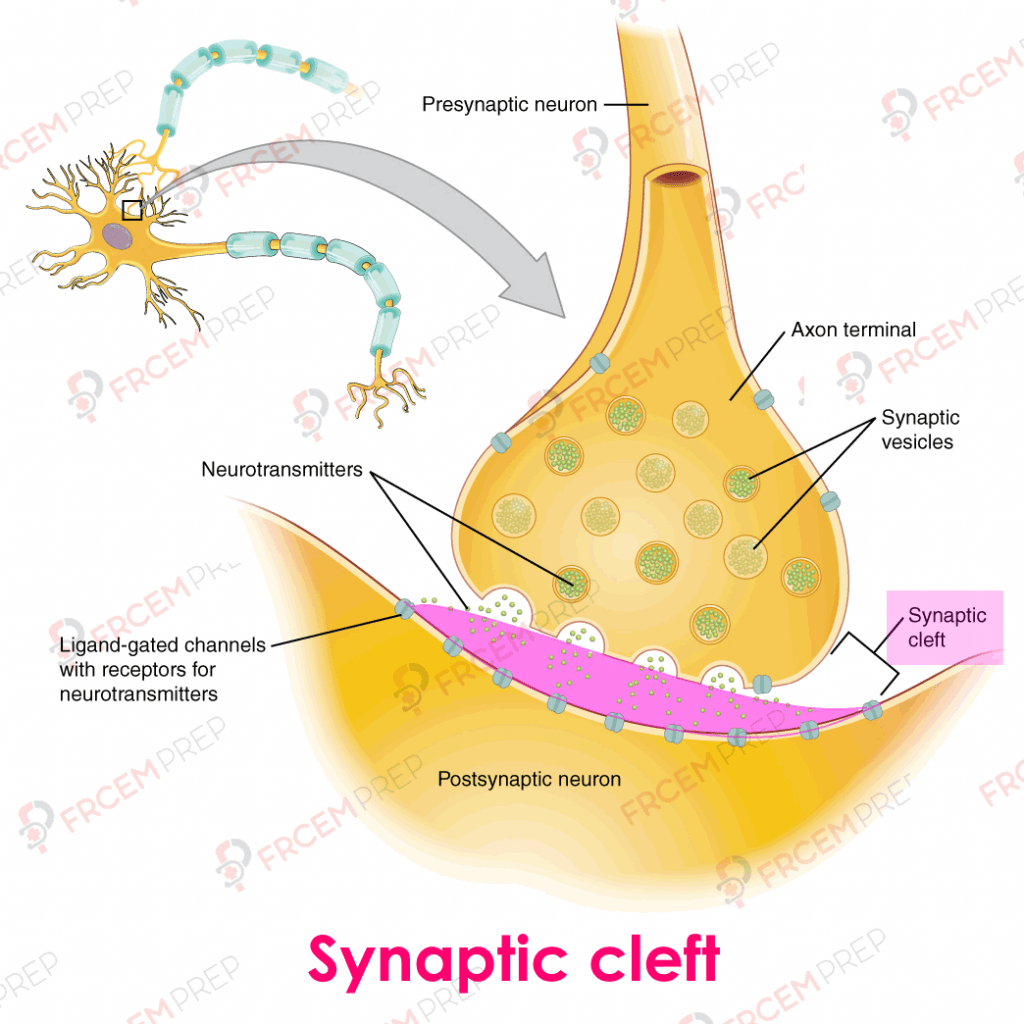
Image modified from wtcs pressbooks.pub CC 4.0
Two neurons communicate at the synapse, where presynaptic bouton is separated from the target by synaptic cleft (Pink area in image).
Narrow cleft = fast and specific signal
Wide cleft = distributed but slower effect.
Synthetic enzymes reach the axon terminal to synthesize neurotransmitter (acetylcholine, noradrenaline), which are stored in vesicles.
Action potential at the nerve ending causes an influx of Ca2+ and vesicles release the neurotransmitter into synapse to act on ligand gated posts-synaptic receptors.
Neurotransmitter release has feedback inhibition via α2-receptors on presynaptic terminal for adrenergic synapses.
Neurotransmitters removal.
In cholinergic synapses, cholinesterase breaks down ACh into choline and acetate for recycling.
In adrenergic synapses, most noradrenaline is rapidly taken up again by the nerve ending via uptake-1 transporter for recycling. Some facilitated diffusion via uptake-2 occurs into smooth muscle.
Excess noradrenaline and sympathomimetic amines such as tyramine (found in some foodstuffs), are metabolized in the neuron by mitochondrial monoamine oxidase (MAO).
Noradrenaline and other catecholamines that enter the circulation are metabolized sequentially by catechol-O-methyl transferase (COMT) and MAO.
If someone is taking MAOI (MAO inhibitors for treating depression), they should not take foods containing excess tyramine (cottage cheese, red wine, smoked fish etc). If taken, can lead to high blood pressure.
Clinical pearl
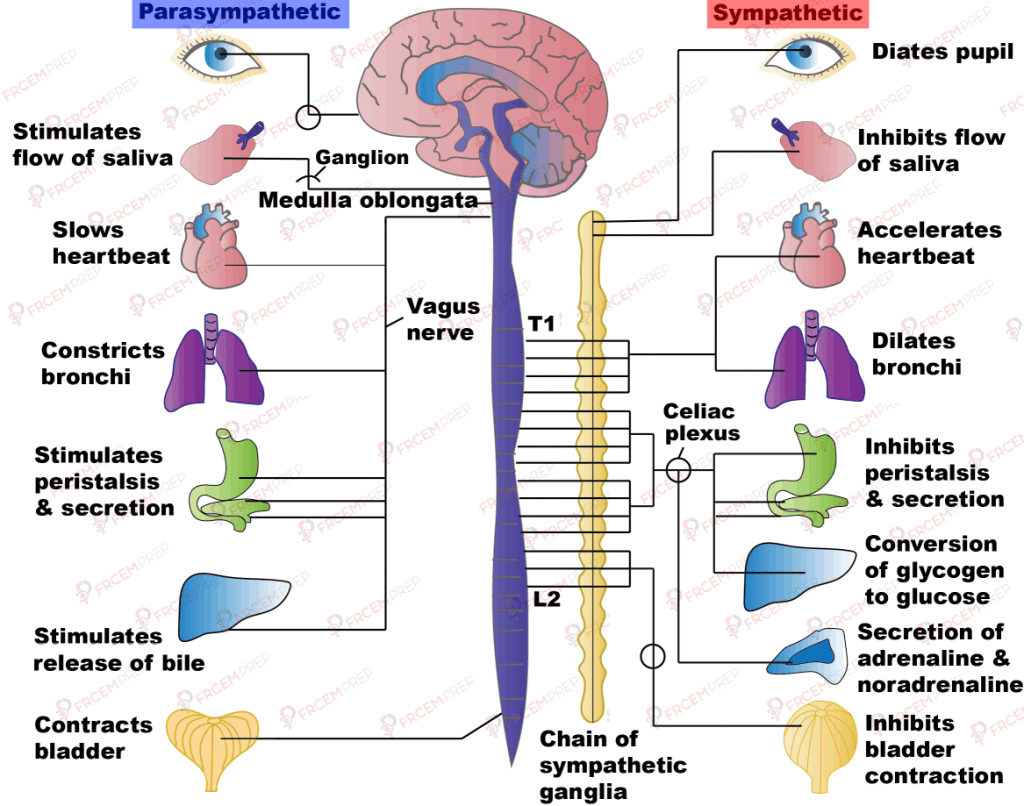
Principal effects of autonomic stimulation
Key effects of SNS & PNS upon body organs
Detail of these effects is covered later in specific cases in relation to systems
Curriculum
Sympathetic system
Preganglionic – from lateral horn of segments T1–L2 of the spinal cord, which exit the cord to reach the ganglia.
Postganglionic neurons release noradrenaline (PNS post ganglionics release ACh).
Noradrenaline and adrenaline – are catecholamines (from Adrenal Medulla).
Adrenergic receptors
Types – α and β, (subdivided into α1, α2, β1, β2, β3).
Noradrenaline and adrenaline are equally potent on α1-receptors [commonly associated with smooth muscle contraction (e.g. blood vessels)].
α2-receptors are mostly inhibitory (On presynaptic nerve terminal to inhibit further release of Neurotransmitter release, and also on blood vessel smooth muscles).
Clonidine is used to control HTN as it causes Vasodilation via α2
Clinical pearl
Noradrenaline is more potent at β1-receptors (Heart rate / force) and adrenaline is more potent at β2-receptors (smooth muscle relaxation at blood vessels and airways).
Non selective beta blockers block beta 1 & 2 equally (Eg. – Propranolol)
Sympathetic neurons releasing acetylcholine at the effector (e.g. sweat glands) – sympathetic cholinergic neurons.
Exception from noradrenaline
Parasympathetic system
Preganglionic neurons start at the brain stem and run in cranial nerves III, VII, IX and X, and also from the S2-S3 sacral segments of spinal cord.
PSN postganglionic neurons release acetylcholine (ACh), [@ cholinergic muscarinic receptors]
Activation increases secretion in many glands (e.g. bronchial mucous glands), and either contraction (e.g. bladder detrusor) or relaxation (e.g. bladder internal sphincter) of smooth muscle.
PSN affects blood vessels only at genitals to help with erection (penis and clitoris), other blood vessels don’t have much response to PSN.